objectifs D’apprentissage
- décrire la structure biochimique des désoxyribonucléotides
- identifier les paires de bases utilisées dans la synthèse des désoxyribonucléotides
- expliquer pourquoi la double hélice de L’ADN est décrite comme antiparallèle
dans le métabolisme microbien, nous avons macromolécules: protéines, lipides et glucides. Dans ce chapitre, nous allons discuter d’une quatrième classe de macromolécules: les acides nucléiques., Comme d’autres macromolécules, les acides nucléiques sont composés de monomères appelés nucléotides, qui sont polymérisés pour former de grosses mèches. Chaque brin d’acide nucléique contient certains nucléotides qui apparaissent dans un certain ordre dans le brin, appelé sa séquence de base. La séquence de base de l’acide désoxyribonucléique (ADN) est responsable du transport et de la conservation de l’information héréditaire dans une cellule., Dans mécanismes de la génétique microbienne, nous discuterons en détail de la façon dont L’ADN utilise sa propre séquence de base pour diriger sa propre synthèse, ainsi que la synthèse de L’ARN et des protéines, ce qui, à son tour, donne lieu à des produits avec une structure et une fonction diverses. Dans cette section, nous allons discuter de la structure et de la fonction de base de L’ADN.
nucléotides D’ADN
Les éléments constitutifs des acides nucléiques sont les nucléotides. Les nucléotides qui composent L’ADN sont appelés désoxyribonucléotides., Les trois composants d’un désoxyribonucléotide sont un sucre à cinq carbones appelé désoxyribose, un groupe phosphate et une base azotée, une structure annulaire contenant de l’azote qui est responsable de l’appariement des bases complémentaires entre les brins d’acide nucléique (Figure 1). Les atomes de carbone de la à cinq atomes de carbone désoxyribose sont numérotées 1ʹ, 2ʹ, 3ʹ, 4ʹ, et 5ʹ (1ʹ est lu comme « une prime”). Un nucléoside comprend le sucre à cinq carbones et la base azotée.

la Figure 1., (a) chaque désoxyribonucléotide est composé d’un sucre appelé désoxyribose, d’un groupe phosphate et d’une base azotée—dans ce cas, l’adénine. b) les cinq carbones contenus dans le désoxyribose sont désignés comme 1ʹ, 2ʹ, 3,, 4ʹ et 5ʹ.
Le désoxyribonucléotide est nommé selon les bases azotées (Figure 2). Les bases azotées adénine (A) et guanine (G) sont les purines; ils ont une structure à double cycle avec un cycle à six carbones fusionné à un cycle à cinq carbones. Les pyrimidines, la cytosine (C) et la thymine (T), sont des bases azotées plus petites qui n’ont qu’une structure à six cycles carbonés.,

la Figure 2. Les bases azotées dans L’ADN sont classées dans les purines à deux anneaux adénine et guanine et les pyrimidines à un anneau cytosine et thymine. La Thymine est unique à L’ADN.
Les triphosphates nucléosidiques individuels se combinent entre eux par des liaisons covalentes appelées liaisons phosphodiester 5ʹ-3ʹ, ou liaisons par lesquelles le groupe phosphate attaché au carbone 5ʹ du sucre d’un nucléotide se lie au groupe hydroxyle du carbone 3ʹ du sucre du nucléotide suivant., La liaison Phosphodiester entre nucléotides forme l’épine dorsale sucre-phosphate, la structure alternée sucre-phosphate composant le cadre d’un brin d’acide nucléique (Figure 3). Au cours du processus de polymérisation, des désoxynucléotides triphosphates (dNTP) sont utilisés. Pour construire l’épine dorsale sucre-phosphate, les deux phosphates terminaux sont libérés du dNTP sous forme de pyrophosphate. Le résultant brin d’acide nucléique est un groupe phosphate à la 5ʹ de carbone et un groupe hydroxyle libre à la 3ʹ carbone fin., Les deux groupes phosphates non utilisés du triphosphate nucléotidique sont libérés sous forme de pyrophosphate lors de la formation de la liaison phosphodiester. Le Pyrophosphate est ensuite hydrolysé, libérant l’énergie utilisée pour conduire la polymérisation des nucléotides.
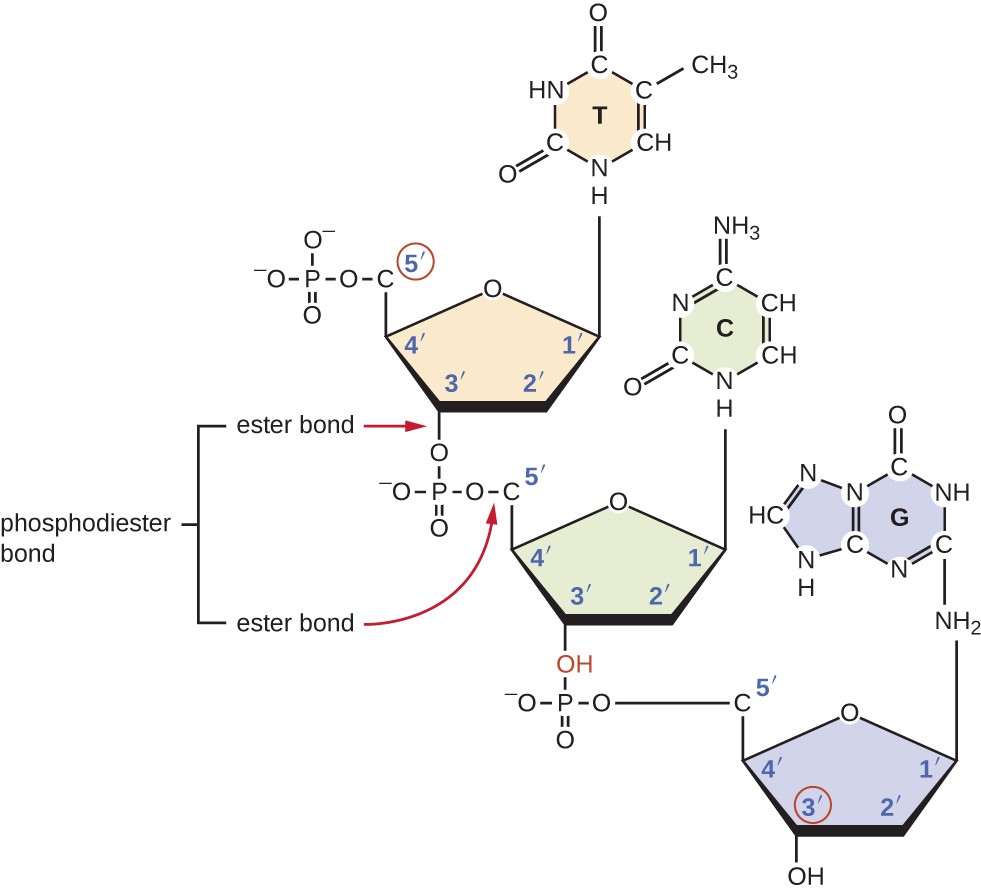
la Figure 3. Des liaisons Phosphodiester se forment entre le groupe phosphate attaché au carbone 5ʹ d’un nucléotide et le groupe hydroxyle du carbone 3ʹ dans le nucléotide suivant, entraînant la polymérisation des nucléotides en brins d’acide nucléique. Notez les extrémités 5ʹ et 3ʹ de ce brin d’acide nucléique.,
pensez-y
- qu’entend-on par les extrémités 5ʹ et 3ʹ d’un brin d’acide nucléique?
découverte de la double hélice
Au début des années 1950, des preuves considérables s’étaient accumulées indiquant que L’ADN était le matériel génétique des cellules, et maintenant la course était lancée pour découvrir sa structure tridimensionnelle., À cette époque, le biochimiste autrichien Erwin Chargaff (1905-2002) a examiné la teneur en ADN de différentes espèces et a découvert que l’adénine, la thymine, la guanine et la cytosine n’étaient pas trouvées en quantités égales et qu’elles variaient d’une espèce à l’autre, mais pas entre les individus de la même espèce. Il a constaté que la quantité d’adénine était très proche d’égaler la quantité de thymine, et la quantité de cytosine était très proche d’égaler la quantité de guanine, ou A = T et G = C. Ces relations sont également connues sous le nom de règles de Chargaff.,

la Figure 4. Le schéma de diffraction des rayons X de L’ADN montre sa nature hélicoïdale. (crédit: National Institutes of Health)
D’autres scientifiques exploraient également activement ce domaine au milieu du 20e siècle. En 1952, le scientifique américain Linus Pauling (1901-1994) était le premier chimiste structural au monde et le favori pour résoudre la structure de l’ADN., Pauling avait auparavant découvert la structure des hélices α des protéines, en utilisant la diffraction des rayons X, et, basé sur des images de diffraction des rayons X de L’ADN faites dans son laboratoire, il a proposé un modèle d’ADN triple brin. Dans le même temps, les chercheurs britanniques Rosalind Franklin (1920-1958) et son étudiant diplômé R. G. Gosling utilisaient également la diffraction des rayons X pour comprendre la structure de l’ADN (Figure 4). C’est L’expertise scientifique de Franklin qui a abouti à la production d’images de diffraction de rayons X plus bien définies de l’ADN qui montreraient clairement la structure globale en double hélice de l’ADN.,
James Watson (1928–), un scientifique américain, et Francis Crick (1916-2004), un scientifique britannique, travaillaient ensemble dans les années 1950 pour découvrir la structure de l’ADN. Ils ont utilisé les règles de Chargaff et les images de diffraction aux rayons X de Franklin et Wilkins des fibres D’ADN pour reconstituer l’appariement purine-pyrimidine de la molécule d’ADN double hélicoïdale (Figure 5). En avril 1953, Watson et Crick ont publié leur modèle de la double hélice D’ADN dans Nature. Le même numéro comprenait en outre des articles de Wilkins et de ses collègues, chacun décrivant différents aspects de la structure moléculaire de l’ADN., En 1962, James Watson, Francis Crick et Maurice Wilkins ont reçu le prix Nobel de physiologie et de Médecine. Malheureusement, à ce moment-là, Franklin était mort et les prix Nobel de l’époque n’étaient pas décernés à titre posthume. Les travaux se sont toutefois poursuivis pour en apprendre davantage sur la structure de l’ADN. En 1973, Alexander Rich (1924-2015) et ses collègues ont pu analyser des cristaux D’ADN pour confirmer et élucider davantage la structure de l’ADN.

la Figure 5., En 1953, James Watson et Francis Crick ont construit ce modèle de la structure de l’ADN, présenté ici au Science Museum de Londres.
pensez-y
- quels scientifiques ont le plus de crédit pour décrire la structure moléculaire de L’ADN?
structure de l’ADN
Watson et Crick ont proposé que l’ADN est composé de deux brins qui sont tordus l’un autour de l’autre pour former une hélice droitière. Les deux brins D’ADN sont antiparallèles, de sorte que l’extrémité 3ʹ d’un brin fait face à l’extrémité 5ʹ de l’autre (Figure 6)., Le 3ʹ fin de chaque brin a un groupe hydroxyle libre, tandis que le 5ʹ fin de chaque brin est un groupe phosphate. Le sucre et le phosphate des nucléotides polymérisés forment l’épine dorsale de la structure, tandis que les bases azotées sont empilées à l’intérieur. Ces bases azotées à l’intérieur de la molécule interagissent les unes avec les autres, l’appariement des bases.
L’analyse des schémas de diffraction de l’ADN a déterminé qu’il y a environ 10 bases par tour dans L’ADN., L’espacement asymétrique des dorsales sucre-phosphate génère des rainures majeures (où l’épine dorsale est éloignée) et des rainures mineures (où l’épine dorsale est rapprochée) (Figure 6). Ces rainures sont des endroits où les protéines peuvent se lier à L’ADN. La liaison de ces protéines peut modifier la structure de l’ADN, réguler la réplication ou réguler la transcription de l’ADN en ARN.

la Figure 6. Watson et Crick ont proposé le modèle à double hélice pour L’ADN., (a) les os dorsaux sucre-phosphate sont à l’extérieur de la double hélice et les purines et les pyrimidines forment les « échelons” de l’échelle de l’hélice D’ADN. (b) les deux brins D’ADN sont antiparallèles l’un à l’autre. (c) la direction de chaque brin est identifiée en numérotant les carbones (1 à 5) dans chaque molécule de sucre. L’extrémité 5ʹ est celle où le carbone # 5 n’est pas lié à un autre nucléotide; l’extrémité 3ʹ est celle où le carbone #3 n’est pas lié à un autre nucléotide.
l’appariement de Base a lieu entre une purine et une pyrimidine., Dans L’ADN, l’adénine (A) et la thymine (T) sont des paires de bases complémentaires, et la cytosine (C) et la guanine (G) sont également des paires de bases complémentaires, expliquant les règles de Chargaff (Figure 7). Les paires de bases sont stabilisées par des liaisons hydrogène; l’adénine et la thymine forment deux liaisons hydrogène entre elles, tandis que la cytosine et la guanine forment trois liaisons hydrogène entre elles.

la Figure 7. Des liaisons hydrogène se forment entre des bases azotées complémentaires à l’intérieur de l’ADN.,
en laboratoire, exposer les deux brins D’ADN de la double hélice à des températures élevées ou à certains produits chimiques peut briser les liaisons hydrogène entre les bases complémentaires, séparant ainsi les brins en deux brins d’ADN simples séparés (ADN simple brin ). Ce processus est appelé dénaturation de l’ADN et est analogue à la dénaturation des protéines, comme décrit dans les protéines. Les brins d’adnss peuvent également être remis ensemble comme ADN double brin (adnsd), par reannealing ou renaturing en refroidissant ou en enlevant les dénaturants chimiques, permettant à ces liaisons hydrogène de se reformer., La capacité de manipuler artificiellement L’ADN de cette manière est à la base de plusieurs techniques importantes en biotechnologie (Figure 8). En raison de la liaison hydrogène supplémentaire entre la paire de bases C = G, L’ADN à haute teneur en GC est plus difficile à dénaturer que l’ADN à faible teneur en GC.

la Figure 8. En laboratoire, la double hélice peut être dénaturée en ADN simple brin par exposition à la chaleur ou à des produits chimiques, puis renaturée par refroidissement ou élimination des dénaturants chimiques pour permettre aux brins d’ADN de se réanneler., (crédit: modification du travail de Hernández-Lemus E, Nicasio-Collazo LA, Castañeda-Priego R)
pensez-y
- quelles sont les deux paires de bases complémentaires de L’ADN et comment sont-elles liées?
fonction ADN
L’ADN stocke les informations nécessaires à la construction et au contrôle de la cellule. La transmission de cette information de la mère aux cellules filles est appelée transfert de gène vertical et elle se produit par le processus de réplication de l’ADN., L’ADN est répliqué lorsqu’une cellule fait une copie en double de son ADN, puis la cellule se divise, ce qui entraîne la distribution correcte d’une copie D’ADN à chaque cellule résultante. L’ADN peut également être dégradé enzymatiquement et utilisé comme source de nucléosides et de nucléotides pour la cellule. Contrairement à d’autres macromolécules, L’ADN ne joue pas de rôle structurel dans les cellules.
pensez-y
- Comment l’ADN transmet-il l’information génétique à la progéniture?,
ouvrir la voie aux femmes dans les sciences et les Professions de la santé
historiquement, les femmes ont été sous-représentées dans les sciences et en médecine, et souvent leurs contributions pionnières sont passées relativement inaperçues. Par exemple, bien que Rosalind Franklin ait réalisé les études de diffraction des rayons X démontrant la structure double hélicoïdale de l’ADN, ce sont Watson et Crick qui sont devenus célèbres pour cette découverte, en s’appuyant sur ses données., Il reste encore une grande controverse quant à savoir si leur acquisition de ses données était appropriée et si les conflits de personnalité et les préjugés sexistes ont contribué à la reconnaissance tardive de ses contributions importantes. De même, Barbara McClintock a fait des travaux pionniers dans la génétique du maïs (maïs) des années 1930 à 1950, découvrant des transposons (gènes sauteurs), mais elle n’a été reconnue que beaucoup plus tard, recevant un prix Nobel de physiologie ou de médecine en 1983 (Figure 9).
Aujourd’hui, les femmes restent encore sous-représentées dans de nombreux domaines de la science et de la médecine., Alors que plus de la moitié des diplômes de premier cycle en sciences sont décernés à des femmes, seulement 46% des doctorats en sciences sont décernés à des femmes. Dans le milieu universitaire, le nombre de femmes à chaque niveau d’avancement professionnel continue de diminuer, les femmes occupant moins du tiers des postes de scientifiques de niveau doctorat dans des postes à durée déterminée, et moins du quart des postes de professeur titulaire dans les collèges et universités de 4 ans., Même dans les professions de la santé, comme presque tous les autres domaines, les femmes sont souvent sous-représentées dans de nombreuses carrières médicales et gagnent beaucoup moins que leurs homologues masculins, comme le montre une étude de 2013 publiée par le Journal of the American Medical Association.
pourquoi de telles disparités continuent-elles d’exister et comment briser ces cycles? La situation est complexe et résulte probablement de la combinaison de divers facteurs, y compris la façon dont la société conditionne les comportements des filles dès leur plus jeune âge et soutient leurs intérêts, à la fois professionnellement et personnellement., Certains ont suggéré que les femmes n’appartiennent pas au laboratoire, y compris le prix Nobel Tim Hunt, dont les commentaires publics de 2015 suggérant que les femmes sont trop émotives pour la science ont été largement condamnés.
Les filles devraient peut-être être davantage soutenues dès leur plus jeune âge dans les domaines des sciences et des mathématiques (Figure 9). Les programmes de science, technologie, ingénierie et mathématiques (STEM) parrainés par L’American Association of University Women (AAUW) et la National Aeronautics and Space Administration (NASA) sont d’excellents exemples de programmes offrant un tel soutien., Les Contributions des femmes à la science devraient être mieux connues du public, et le marketing destiné aux jeunes filles devrait inclure davantage d’images des femmes scientifiques et professionnelles ayant réussi historiquement et professionnellement, encourageant tous les jeunes esprits brillants, y compris les filles et les femmes, à poursuivre une carrière dans les sciences et la médecine.

la Figure 9. (a) les travaux de Barbara McClintock sur la génétique du maïs dans les années 1930 à 1950 ont abouti à la découverte de transposons, mais son importance n’a pas été reconnue à l’époque., b) les Efforts visant à encadrer de manière appropriée les femmes dans les domaines de la science et de la médecine et à leur apporter un soutien social continu pourraient un jour contribuer à atténuer certains des problèmes qui empêchent l & apos; égalité des sexes à tous les niveaux dans les domaines de la science et de la médecine. (crédit a: modification du travail par la Smithsonian Institution; crédit b: modification du travail par Haynie SL, Hinkle AS, Jones NL, Martin CA, Olsiewski PJ, Roberts MF)
Focus clinique: Aamir, Partie 2
cet exemple poursuit L’histoire D’Aamir qui a commencé en utilisant la microbiologie pour découvrir les Secrets de la vie.,
sur la base de ses symptômes, le médecin d’Aamir soupçonne qu’il souffre d’une maladie d’origine alimentaire qu’il a contractée lors de ses voyages. Les possibilités comprennent une infection bactérienne (par exemple, E. coli entérotoxigène, Vibrio cholerae, Campylobacter jejuni, Salmonella), une infection virale (rotavirus ou norovirus) ou une infection protozoaire (Giardia lamblia, Cryptosporidium parvum ou Entamoeba histolytica).
son médecin ordonne un échantillon de selles pour identifier les agents responsables possibles (p. ex., bactéries, kystes) et pour rechercher la présence de sang parce que certains types D’agents infectieux (comme C., jejuni, Salmonella et E. histolytica) sont associés à la production de selles sanglantes.
L’échantillon de selles D’Aamir ne montrait ni sang ni kystes. À la suite de l’analyse de son échantillon de selles et d’après ses récents antécédents de voyage, le médecin de l’hôpital a soupçonné Qu’Aamir souffrait de diarrhée du voyageur causée par E. coli entérotoxigène (ETEC), l’agent causal de la plupart des diarrhées du voyageur., Pour vérifier le diagnostic et exclure d’autres possibilités, le médecin D’Aamir a ordonné un test de laboratoire de diagnostic de son échantillon de selles pour rechercher des séquences D’ADN codant des facteurs de virulence spécifiques d’ETEC. Le médecin a demandé à Aamir de boire beaucoup de liquides pour remplacer ce qu’il perdait et l’a renvoyé de l’hôpital.
ETEC produit plusieurs facteurs de virulence codés par plasmides qui le rendent pathogène par rapport à E. coli typique. Ceux-ci comprennent les toxines sécrétées chaleur-entérotoxine labile (LT) et chaleur-entérotoxine stabile (ST), ainsi que le facteur de colonisation (CF)., LT et ST provoquent l’excrétion des ions chlorure des cellules intestinales vers la lumière intestinale, entraînant une perte conséquente d’eau des cellules intestinales, entraînant une diarrhée. CF code une protéine bactérienne qui aide à permettre à la bactérie d’adhérer à la muqueuse de l’intestin grêle.
- pourquoi le médecin D’Aamir a-t-il utilisé l’analyse génétique au lieu d’isoler les bactéries de l’échantillon de selles ou de colorer directement L’échantillon de selles?
Nous reviendrons à L’exemple D’Aamir dans les pages suivantes.,
Concepts clés et résumé
- Les acides nucléiques sont composés de nucléotides, chacun contenant un sucre pentose, un groupe phosphate et une base azotée. Les désoxyribonucléotides dans L’ADN contiennent du désoxyribose comme sucre pentose.
- L’ADN contient les pyrimidines cytosine et thymine, et les purines adénine et guanine.
- Les nucléotides sont reliés entre eux par des liaisons phosphodiester entre le groupe 5 phosphate phosphate d’un nucléotide et le groupe 3 Hydrox hydroxyle d’un autre., Un brin d’acide nucléique est un groupe phosphate à la 5ʹ fin et un groupe hydroxyle libre à la 3ʹ fin.
- Chargaff a découvert que la quantité d’adénine est approximativement égale à la quantité de thymine dans L’ADN, et que la quantité de guanine est approximativement égale à la cytosine. Ces relations ont ensuite été déterminées comme étant dues à l’appariement de base complémentaire.
- Watson et Crick, s’appuyant sur les travaux de Chargaff, Franklin et Gosling, et Wilkins, ont proposé le modèle de double hélice et l’appariement de base pour la structure de l’ADN.,
- l’ADN est composé de deux brins complémentaires orientés antiparallèle à l’autre avec les squelettes phosphodiester à l’extérieur de la molécule. Les bases azotées de chaque brin se font face et les bases complémentaires se lient l’une à l’autre, stabilisant la double hélice.
- La chaleur ou les produits chimiques peuvent briser les liaisons hydrogène entre les bases complémentaires, dénaturant L’ADN. Le refroidissement ou l’élimination des produits chimiques peut conduire à la renaturation ou à la réannuration de l’ADN en permettant aux liaisons hydrogène de se reformer entre les bases complémentaires.,
- L’ADN stocke les instructions nécessaires pour construire et contrôler la cellule. Cette information est transmise du parent à la progéniture par transfert de gène vertical.
Plusieurs Choix
Lequel des éléments suivants n’est pas trouvé dans l’ADN?
- la thymine
- phosphodiester liens
- appariement de bases complémentaires
- acides aminés
Si 30% des bases d’une molécule D’ADN sont de l’adénine, Quel est le pourcentage de thymine?
- 20%
- 25%
- 30%
- 35%
laquelle des déclarations suivantes sur l’appariement de base dans L’ADN est incorrecte?
- Les Purines sont toujours paires de base avec les pyrimidines.
- l’Adénine se lie à la guanine.
- les paires de bases sont stabilisées par des liaisons hydrogène.,
- Base de couplage se produit à l’intérieur de la double hélice.
Si un brin D’ADN contient la séquence 5ʹ-ATTCCGGATCGA-3ʹ, laquelle des séquences suivantes est la séquence du brin complémentaire de L’ADN?
- 5ʹ-TAAGGCCTAGCT-3ʹ
- 5ʹ-ATTCCGGATCGA-3ʹ
- 3ʹ-TAACCGGTACGT-5ʹ
- 5ʹ-TCGATCCGGAAT-3ʹ
Au cours de la dénaturation de L’ADN, lequel des événements suivants se produit?
- Les liaisons hydrogène entre bases complémentaires se brisent.
- Les liaisons Phosphodiester se brisent dans l’épine dorsale sucre-phosphate.
- Les liaisons hydrogène dans l’épine dorsale sucre-phosphate se brisent.
- Les liaisons Phosphodiester entre bases complémentaires se brisent.
à compléter
La fin d’un brin d’acide nucléique avec un gratuit groupe phosphate est appelé ________.
Vrai/Faux
Les travaux de Rosalind Franklin et R. G. Gosling ont été importants pour démontrer la nature hélicoïdale de L’ADN.,
La paire de base a plus de liaison hydrogène de la C-G paire de base.
Penser
- Quel est le rôle de phosphodiester obligations dans le squelette sucre-phosphate de l’ADN?
- qu’entend-on par le terme « antiparallèle? »
- pourquoi L’ADN avec une teneur élevée en GC est-il plus difficile à dénaturer que celui avec une faible teneur en GC?,
- En considérant la structure de la double hélice D’ADN, comment vous attendez-vous à ce que la structure diffère s’il y avait appariement de base entre deux purines? Entre deux pyrimidines?
- Un certain échantillon D’ADN a une composition composée de 22% de thymine. Utilisation Chargaff règles de remplir les pourcentages pour les trois autres bases azotées.,
| base | adenine | guanine | thymine | cytosine |
|---|---|---|---|---|
| % | 22% |